A Multi-Megabase Copy Number Gain Causes Maternal Transmission Ratio Distortion on Mouse Chromosome 2
Abstract:
Significant departures from expected Mendelian inheritance ratios (transmission ratio distortion, TRD) are frequently
observed in both experimental crosses and natural populations. TRD on mouse Chromosome (Chr) 2 has been reported in multiple
experimental crosses, including the Collaborative Cross (CC). Among the eight CC founder inbred strains, we found that Chr 2
TRD was exclusive to females that were heterozygous for the WSB/EiJ allele within a 9.3 Mb region (Chr 2 76.9 - 86.2 Mb).
A copy number gain of a 127 kb-long DNA segment (designated as responder to drive, R2d) emerged as the strongest candidate
for the causative allele. We mapped R2d sequences to two loci within the candidate interval. R2d1 is located near the proximal
boundary, and contains a single copy of R2d in all strains tested. R2d2 maps to a 900 kb interval, and the number of R2d copies
varies from zero in classical strains (including the mouse reference genome) to more than 30 in wild-derived strains. Using
real-time PCR assays for the copy number, we identified a mutation (R2d2WSBdel1) that eliminates the majority of
the R2d2WSB
copies without apparent alterations of the surrounding WSB/EiJ haplotype. In a three-generation pedigree segregating for
R2d2WSBdel1, the mutation is transmitted to the progeny and Mendelian segregation is restored in females heterozygous for
R2d2WSBdel1, thus providing direct evidence that the copy number gain is causal for maternal TRD. We found that transmission
ratios in R2d2WSB heterozygous females vary between Mendelian segregation and complete distortion depending on the genetic
background, and that TRD is under genetic control of unlinked distorter loci. Although the R2d2WSB transmission ratio was
inversely correlated with average litter size, several independent lines of evidence support the contention that female
meiotic drive is the cause of the distortion. We discuss the implications and potential applications of this novel
meiotic drive system.
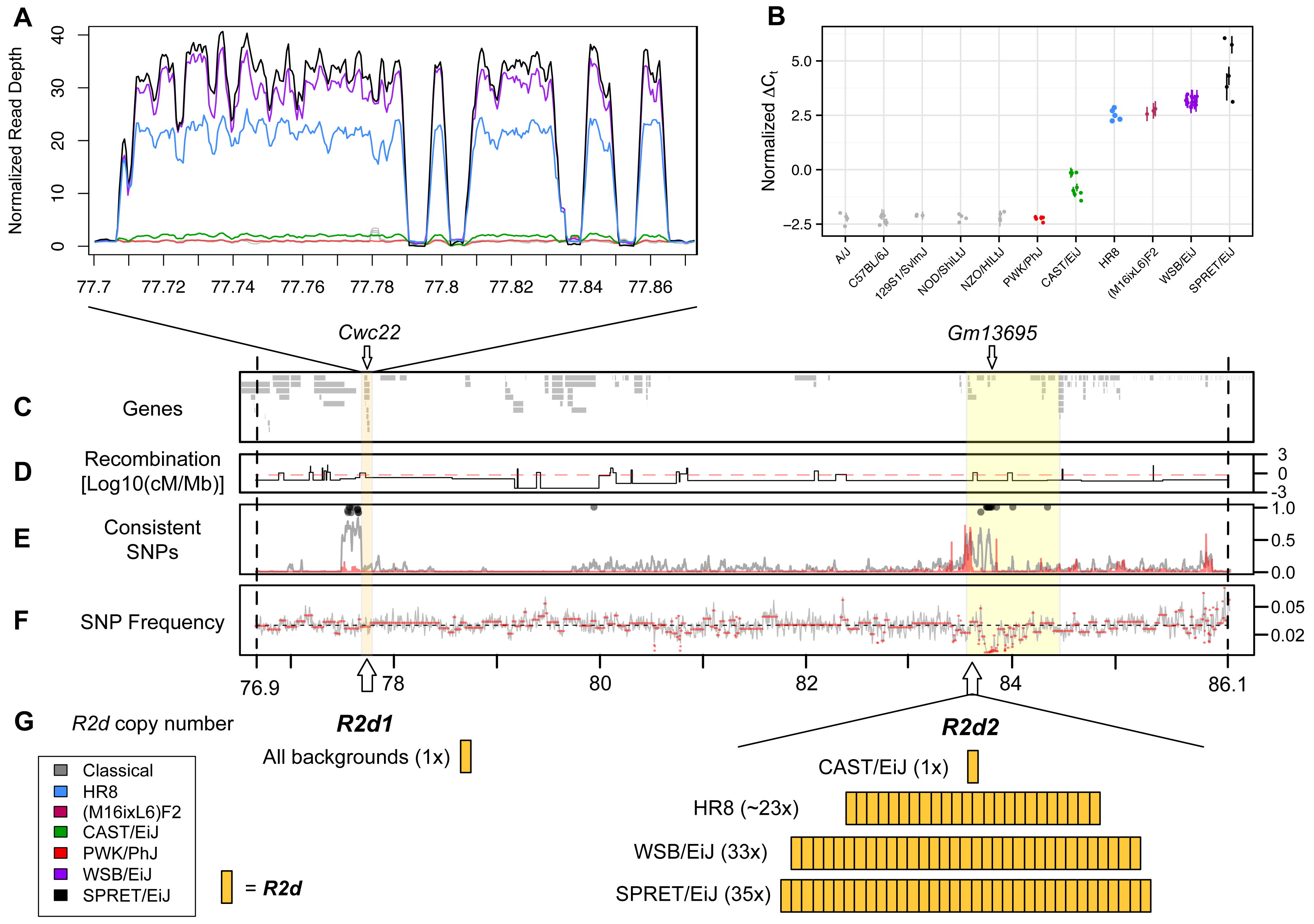
Figure 2. A large copy number gain is present in strains with maternal TRD. A) Read depth in 100 bp windows, normalized by the
genome-wide mean read depth for each strain, for R2d1, a 158 kb region within the 9.3 Mb candidate interval defined in Fig. 1.
R2d1 includes a single (non-contiguous) copy of R2d. Strains are represented by the colors shown in the inset. Inbred strains
included under the heading "classical" are A/J, 129S1/SvImJ, C57BL/6JN, NOD/ShiLtJ and NZO/HILtJ. The four large gaps represent LINEs that
were inserted in the unique copy found in the reference genome after initial duplication. B) R2d copy number estimated by TaqMan assays
for Cwc22. Normalized cycle threshold (ΔCt; see Methods) is proportional to absolute copy number on the log scale. Strains are colored
as in panel A. The (M16ixL6)F2 samples shown are known to be homozygous for the M16i allele based on genotypes from the MegaMUGA array.
C-F) The yellow boxes highlight the 158 kb region depicted in panel A (R2d1) and the 900 kb R2d2 locus mapping interval.
Vertical dashed lines indicate the boundaries of the 9.3 Mb candidate interval. C) Locations of Ensembl genes in the NCBI/37 reference
genome within the interval. The locations of the Cwc22 gene and of seven Cwc22 pseudogenes (Gm13695), are shown.
D) Recombination frequency based on Liu et al. (2014), normalized by physical distance (Mb) and log10-transformed. The red line indicates
the mean recombination frequency for Chr 2. E) Frequency distribution measured in 1 kb windows of SNPs with shared alleles among the three
strains with TRD (WSB/EiJ, SPRET/EiJ and HR8; gray line), and with alleles perfectly consistent between strains with TRD and strains without
TRD (A/J, 129S1/SvImJ, C57BL/6JN, NOD/ShiLtJ, NZO/HILt, CAST/EiJ and PWK/PhJ; red line). Lines are smoothed. Black circles indicate windows
in which the strains with TRD share an allele for at least 90% of SNPs. F) Frequency distribution of reported SNPs in the candidate interval.
G) The location and the number of copies of R2d that are present in R2d1 and R2d2.